| Иван Ефремов - Тафономия и геологическая летопись Страницы: 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 51, 52, 53, 54, Глава VII. Примеры тафономических поправок к вопросам эволюции наземных позвоночных
В предыдущем изложении мы видели, что совокупность данных геологической летописи создалась не случайным путем. Какое же значение и какое влияние на наши представления о жизни древних геологических эпох могут иметь тафономические закономерности?
Прежде всего нужно отметить, что закономерная неполнота наших представлений о наземной жизни далекого прошлого является как бы односторонней. Из всей суммы разнообразия древних наземных форм выборочно выпадают все "малые числа", т. е. формы, представленные малым числом индивидов, редкие или неплотно заселяющие ареал своего обитания. Нетрудно видеть, что все возможные "переходные" формы как раз попадают под уничтожение в закономерностях образования геологической летописи. Если представить себе какую-нибудь фауну наземных позвопочных, обитающую в области, близкой к области осадконакопления, то составляющие ее формы при переходе в геологическую летопись расположатся по следующим основным группам.
1) Преобладающие количественно в данный момент формы водных или близких к водному обитанию адаптации. Эти формы при достаточной плотности населения перейдут в геологическую летопись.
2) Формы, наиболее прогрессивные, в начале адаптивной радиации завоевавшие новые ареалы обитания и поэтому либо немногочисленные, либо сильно удаляющиеся от области захоронения. Эти формы выпадут из геологической летописи, по крайней мере в подавляющем числе.
3) Формы архаические, уже сошедшие со сцены и вымирающие группы и поэтому редкие. Эти формы также выпадут из геологической летописи, но для очень древних эпох наземной жизни, как мы увидим ниже, имеются исключения.
Таким образом, при переходе в геологическую летопись каждая определенная наземная фауна распадается на три группы форм, из которых в огромном большинстве случаев захороняется только одна. Эта группа форм, достигших большого количественного расцвета, будет состоять из зачастую различных по своей эволюционной стадии форм, с ярко выраженной специализацией, которая определила их благополучие в естественном отборе на данный момент и, следовательно, их количественное преобладание в фауне. Кроме того, это формы, способные населять ареал своего распространения со значительной плотностью или же периодически образовывать большие скопления индивидов — стада. Характер адаптации захороняющейся группы форм будет также подобранным в смысле преобладания адаптации к обитанию в прибрежных областях или вообще областях крупного осадконакопления (рис. 42).
Таким образом, отсутствие в геологической летописи форм, обладающих переходным между крупными и мелкими группами строением, вполне закономерно и отнюдь не может считаться за действительное их отсутствие[1]. Выпадение из геологической летописи особенно интересных по своему эволюционному значению форм, как и вообще выпадение значительного количества форм, слагающих многообразие наземного животного мира любой древней геологической эпохи, определяет своеобразные искажения эволюционных представлений.
Представим себе в общих чертах геологическую документацию смены фауны в эволюции позвоночных от палеозоя до наших дней.
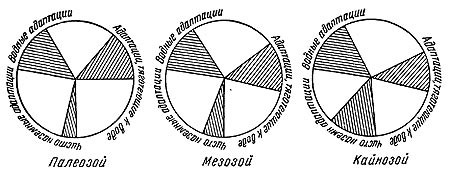
Рис. 42. Схема соотношения типов адаптации наземных форм, представленных в местонахождениях разных геологических эр. Заштрихована сохраняющаяся в геологической летописи часть. Для кайнозоя показано примерно равное соотношение водных и наземных адаптации, исходя из соображения, что эти адаптации частью захоронены в еще не вскрытых ультрафациях и, таким образом, находятся в летописи.
В поясе ультрафаций, сохраняющемся во времени от материка, вся активно эволюционирующая, наиболее прогрессивная часть фауны будет находиться на внутреннем (для наземных) или внешнем (для морских) контуре пояса. В пределах самого пояса, в слагающих его ультрафациях будут захороняться наиболее многочисленные, давно сложившиеся и составленные уже сильно специализованными формами биоценозы.
В ультрафациях силура и нижнего девона мы будем находить сильно специализированных рыб, какими являются известные панцирные рыбы. Родоначальные или архаические формы этих рыб, не обладающие панцирем или окостеневшим внутренним скелетом, помимо своей редкости не будут сохраняться еще и по условиям фоссилизации. Другие формы рыб, несомненно существовавшие одновременно с панцирными, но эволюционировавшие в направлении завоевания моря, рассеются на внешнем контуре зоны захоронения, не образуя достаточных концентраций или вследствие недостаточной индивидуальной численности, или вследствие подвижного образа жизни вне основных зон осадконакопления. Примером подобного выпадения из геологической летописи крупных рыб, обитающих на внешнем контуре пояса захоронения, является Latimeria. Как известно, это огромная кистеперая рыба из семейства Undinidae была совсем недавно поймана у берегов Южной Африки. Последние представители кистеперых сем. Undinidae вымерли в верхнем мелу по данным геологической летописи и, однако, Latimeria оказалась дожившей до наших дней. Случай с Latimeria показывает, что в оценку эволюционной ступени и общего характера фауны древнейших рыб должны быть введены существенные поправки на выпавшие из летописи группы форм подвижных или немногочисленных в отношении количества особей, как раз составляющих наиболее прогреет сивную часть фауны с предковыми формами наземных позвоночных. В поясах материковых ультрафаций верхнего палеозоя внутренний кон-тур пояса подвинется дальше в материк, захватывая область обитания наиболее многочисленной наземной фауны в дельтах, нижнем течении крупных рек. Для каждого из периодов геологической летописи, за исключением разве самых позднейших, характерно отсечение и исчезновение из циклов захоронения всех далеко проникающих в глубь материка филогенетических ветвей.
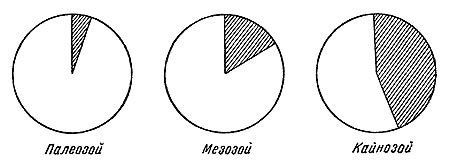
Рис. 43. Схема соотношения захороненной в геологической летописи части животного мира наземных позвоночных (то же для растений) к действительно существовавшей для разных эр геологической истории.
В мезозое и кайнозое круг попадающих в захоронение наземных форм внутреннего контура соответственно расширится, в особенности в кайнозое, благодаря сохранению во времени таких континентальных отложений, которые для более древних эпох являются инфрафациями. Соответственно этому для мезозоя и кайнозоя возрастает многообразие наземных форм, что и кажется вполне естественным как постепенное завоевание суши наземными позвоночными. Однако эта постепенность завоевания, якобы связанная с общим прогрессом, будет в значительной мере только кажущейся, получаясь в результате общих закономерностей сохранения осадков во времени. В самом деле, уже в эоцене мы наблюдаем появление всех главных типов млекопитающих. Более древние жизненные формы, как, например, ящерицы и змеи, обитают в самых различных внутриматериковых районах современности, а современные роды амфибий известны уже со среднего эоцена (верхнелютетские угли Гейзельталя). Нельзя не обратить внимания на тот факт, что, чем больше форм становится нам известно в геологической летописи, тем глубже уходят во время "корни"— исходные формы, не только целых классов, но и отдельных больших групп. Все эти явления могут быть суммированы в следующем выводе: широкое заселение материков и разнообразие животной и растительной жизни на суше возникли значительно ранее, чем это представляется нам по данным геологической летописи, в более древних страницах которой исчезает все большая часть наземного животного мира (рис. 43). Проведение исторической грани между первичным завоеванием суши и кажущейся "пустотой" древних материков — одна из важнейших задач палеонтологии в ее будущем развитии.
Периодические изменения хода геологических процессов также своеобразно отражаются на общей картине геологической документации наземной жизни. Смена периодов покоя и орогенеза неминуемо вносит изменения в совокупность процессов образования местонахождений. В периоды покоя, при низком уровне материков, медленной эрозии и медленном накоплении осадков пояса ультрафаций с захороненной в них фауной будут относительно невелики по абсолютной мощности, располагаются ближе к материку и имеют несколько большую ширину вследствие того, что низменные прибрежные области будут глубоко вдаваться в материк. Процессы захоронения в периоды покоя будут значительно более медленными в соответствии с медленностью осадконакопления, что вызовет большую деструкцию захороненных остатков. Поэтому общее количество, в особенности для внутриматериковых областей, наземной фауны, захороненной в период покоя, будет относительно малым, с меньшим числом местонахождений на данный объем вмещающей породы.
Периоды диастрофизма результируются в энергичной эрозии и увеличении скорости осадконакопления. Благодаря быстрому сносу и захоронению остатков, несмотря на более многочисленные внутриформационные перемывы, количество захороненной фауны и число местонахождений относительно больше. Равным об разом, больше и число форм чисто наземных адаптации благодаря быстрому сносу из внутренних частей материка. Пояса окраинных ультрафаций значительны по мощности и дальше выдаются в море от материка. Субаэральные части дельт в периоды диастрофизма, как правило, относительно уже (рис. 44.).
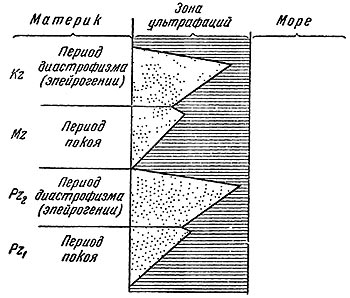
Рис. 44. Диаграмма захоронения в периоды диастрофизма и покоя.
1 Подразумеваются "переходные" формы геологической летописи, как правило — таксономические единицы крупнее вида, отнюдь не исключающие разрыва (скачка) в филогенетическом ряду.
|