| Иван Ефремов - Тафономия и геологическая летопись Страницы: 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 51, 52, 53, 54,
Условия образования концентрации животных остатков довольно разнообразны, но, как правило, исходной точкой всех этих условий служит большая численность индивидов в той или иной местности. Из этого явствует, что значительные концентрации остатков могут получиться только в фаунах, достигших большого количественного расцвета, фаунах преуспевающих или мигрирующих. В составе любой фауны мы встречаем формы с различной численностью индивидов, есть формы весьма редкие и формы, весьма часто встречающиеся. Создавшаяся в фауне концентрация животных остатков будет составлена из форм с большой численностью особей, могущих дать такое скопление. Формы редкие, с малой численностью индивидов, концентраций не дадут и выпадут из захоронения с самых первых шагов образования местонахождения. Таким образом, выборочность захоронения начинается с первого же подготовительного, так сказать, процесса образования скоплений остатков фауны. Фауны, разнообразные по своему составу, но немногочисленные по количеству особей, могут совсем не дать концентраций животных остатков, достаточных для образования местонахождений, и, следовательно, выпасть из геологической летописи.
Вторым фактором образования концентраций животных остатков, сопряженным с общей численностью индивидов, является плотность населения. Повышенная плотность есть благоприятный фактор для образования скоплений остатков. Редкое население может не попадать в захоронение несмотря на длительную заселенность какой-нибудь большой площади. Как известно, формы плодовитые и недолговечные отличаются повышенной плотностью населения и, следовательно, могут преимущественно попадать в захоронение. Формы долговечные и малоплодовитые имеют малую плотность населения и, попадая в захоронение с гораздо большим трудом, определяют еще одну возможность искажения подлинного состава фауны в местонахождении. В основе образования концентраций животных остатков для свободно передвигающихся наземных форм, остатки которых обычно рассеиваются по субаэральной поверхности материка, лежат две главные причины с многочисленными вариациями, в зависимости от бесконечного разнообразия процессов биосферы. Первая причина — массовая гибель животных, как мы увидим далее, отнюдь не связана с катастрофами в виде резких геологических переворотов, катаклизмов, как это представлялось ранее и, зачастую, в скрытом виде отражено в современных представлениях. Катастрофы, несущие массовую гибель животных, в подавляющем большинстве случаев являются таковыми только для определенных биоценозов или областей, населенных фауной, и отнюдь не имеют повсеместного, универсального значения. К явлениям этого типа принадлежат, например, эпизоотии, периодические наводнения в низменных областях, высыхание водоемов, засасывающие болота, неожиданные снеговые бури и так далее. Зачастую стечение обстоятельств, гибельное для некоторых форм определенного биоценоза, совершенно не затрагивает остальных членов этого биоценоза. В качестве примера можно привести колоссальную эпизоотию антилоп в Восточной и Южной Африке, уничтожившую во второй половине прошлого века многие миллионы особей, но совсем не затронувшую носорогов, слонов и других членов биоценоза саванн Африки (Elton 1931). Затопления в зимние периоды низменных областей бассейна Амазонки и Ориноко в Южной Америке, или Лимпопо и Конго в Африке несут массовую гибель мелким членам фауны или молодняку, не трогая крупных форм. Подобных примеров избирательного значения катастроф массовой гибели наземных животных можно привести очень много (Weigelt 1927; Zelicko 1936). Для биоценозов с высокой численностью особей периодическое наступление массовой гибели, вследствие голода и эпизоотии, вообще закономерно и является одним из факторов, регулирующих биотическое равновесие (Nicholson 1933; Северцов 1941). Второй причиной создания концентраций остатков наземной фауны является постепенное накопление остатков, в случае долговременного существования соответствующих условий, в озерах, болотах или дельтах рек, куда в течение очень длительных промежутков времени могут приноситься остатки животных. Постепенное накопление остатков (в количестве, достаточном для образования местонахождения), вообще, встречается реже массовой гибели, но на протяжении длительных геологических периодов все же часто имело место. Нетрудно видеть, что количество животных остатков, нужное для образования местонахождения в инфрафации, может быть значительно меньшим, чем в ультрафации. Действительно, для верхнего кайнозоя и четвертичного периода мы имеем очень большое число таких малых местонахождений. Поэтому в более молодых геологических периодах постепенное накопление животных остатков приобретает все большее значение, так как все меньшее количество остатков требуется для образования заметных нам местонахождений в мелких инфрафациях (рис. 28).
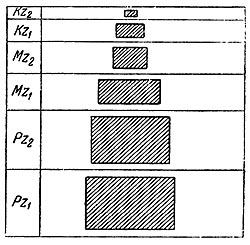
Рис. 28. Диаграмма относительных размеров скоплений органических остатков, необходимых для образования местонахождений в разные геологические эпохи.
Каковы бы ни были причины, лежащие в основе образования скоплений животных остатков, и как велики бы ни были подобные концентрации, деструкция органических остатков будет неизбежной, если данная концентрация не находится в области крупной седиментации или поблизости от нее. Если скопление животных остатков образовалось в пределах седиментационной обстановки ультрафации или, по крайней мере, это скопление может быть вынесено туда достаточно быстро, чтобы избежать деструкции при выветривании и деструкции при переносе, — начинается второй этап образования местонахождений (рис. 29). Нельзя забывать, что всякое местонахождение наземных форм образовано в условиях, заведомо чуждых обстановке жизни данных форм. Все без исключения местонахождения наземной фауны (и флоры) являются, по старой терминологии, танатоценозами (Wasmund 1926). Поэтому всякое скопление животных остатков, подготовленное процессами биосферы к захоронению, включает как необходимое понятие — смерть. Смерть является тем фактором, который в случае наличия концентрации живых животных в данном месте фиксирует скопление, завершая первый этап образования местонахождений.
Для форм водной адаптации, могущих обитать непосредственно в обстановке седиментации, смерть является фактором, как бы оставляющим их в этой обстановке и исключающим возможность ухода из области захоронения. Поэтому концентрации водных форм не нуждаются в обязательном, для вполне наземных, факторе выноса остатков в седиментационную обстановку. Однако все остальные условия остаются в силе и в этом случае. По указанным обстоятельствам случаи постепенного накопления остатков для водных форм могут встречаться гораздо более часто. По тем же причинам, представители водных или тяготеющих к воде адаптации животного мира материков должны значительно чаще попадать в захоронение и чаще образовывать местонахождения.
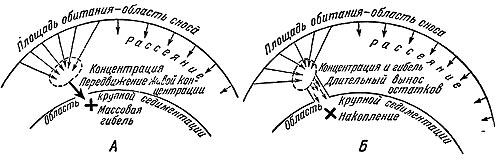
Рис. 29. Биологические факторы образования местонахождений.
А — танатоценоз и тафоценоз при массовой гибели; Б — то же при постепенном накоплении остатков.
Биологические факторы образования местонахождений наземной флоры имеют несколько иной характер. Все общие предпосылки рассеяния остатков и необходимости большой численности и плотности населения имеют ту же силу. Однако концентрации живых растительных форм неподвижны и поэтому возможность попадания массовых и полных остатков в седиментационные условия ультрафаций ограничена. Если растительная концентрация сама находится в ультрафации, то из нее может образоваться мощное местонахождение, гораздо более полно отражающее местный состав флоры, чем мы это имеем в танатоценозах захороненной фауны. В этом отношении условия захоронения флоры приближаются к условиям захоронения морской фауны. Однако скопления флоры вне седиментационных условий ультрафаций, в инфрафациях, доходят оттуда в ультрафации в сильно поврежденном и неполном состоянии, поскольку возможности случайного передвижения в биосфере к областям ультрафаций исключены для растений. Поэтому материковая флора из областей инфрафаций известна в геологической летописи значительно хуже фауны, с гораздо меньшей полнотой остатков, в особенности для палеозоя и мезозоя. В верхнем мезозое и кайнозое при очень большом сходстве с современной флорой, даже по незначительным остаткам, имеется возможность представить себе общий облик растения, почему и получается кажущаяся полнота летописи.
Изложенные выше биологические факторы образования местонахождений наземной фауны, разумеется, не могут претендовать даже на приблизительную полноту. Задачей нашего исследования было показать общий план процессов, действующих со стороны биосферы, и тем самым наметить пути для дальнейших исследований в этой области. Некоторые важнейшие следствия основных биологических факторов будут очерчены нами в разделе общих выводов о факторах образования местонахождений.
2. Главные факторы процессов захоронения
Второй этап образования местонахождений — захоронение накапливающихся органических остатков — в сущности совпадает с процессом седиментации. Преобразования умерших животных и растительных организмов на пути к месту отложения осадка составляют основу влияния условий захоронения на состав отлагавшейся в местонахождении фауны и, следовательно, на полноту геологической летописи. В процессе захоронения органические остатки являются частью отлагаемого осадка и подчиняются всем закономерностям седиментации кластических пород. Поэтому изучение пород, слагающих местонахождение, равно как и общих генетических данных костеносной толщи в целом, дает нам факты, позволяющие восстановить наиболее характерные черты данного захоронения. Сопряженное изучение распределения остатков внутри пласта, размеров и характера истирания органических остатков, короче говоря, совокупность черт состояния остатков флоры и фауны в местонахождении до фоссилизации, вскрывает характер преобразований первичного скопления животных остатков (танатоценоза) и, вместе с данными изучения пород местонахождения, дает картину генезиса "фауны", т. е. тафоценоза (Quenstedt 1931), остатки которой захоронены в местонахождении.
|